Home > A. Molecular pathology > serpins
serpins
Wednesday 29 October 2003
Definition: Serpins are a large serine protease inhibitor gene family.
The serpin superfamily of serine proteinase inhibitors has a central role in controlling proteinases in many biological pathways in a wide range of species.
Serpins are a group of proteins with similar structures that were first identified as a set of proteins able to inhibit proteases. The name serpin is derived from this activity as serine protease inhibitors. They have a conserved structure that have been identified in nearly all species and act as suicide substrates by binding covalently to their target proteases. Serpins regulate various physiological processes and defence mechanisms.
The inhibitory function of the serpins involves a marked conformational transition, but this inherent molecular flexibility renders the serpins susceptible to point mutations that result in aberrant intermolecular linkage and polymer formation (see serpinopathies). The effects of such protein aggregation are cumulative, with a progressive loss of cellular function that results in diseases as diverse as cirrhosis and pulmonary emphysema, as in alpha1-antitrypsin deficiency.
Over 1000 serpins have now been identified, these include 36 human proteins, as well as molecules in plants, bacteria, archaea and certain viruses. Serpins are thus the largest and most diverse family of protease inhibitors.
Serpins undergo a unique and dramatic change in shape (or conformational change) when they inhibit target proteases. This is unusual because most classical protease inhibitors function as simple "lock and key" molecules that bind to and block access to the protease active site (see for example, bovine pancreatic trypsin inhibitor).
Examples of serpins
![]() corticosteroid-binding globulin
corticosteroid-binding globulin![]() alpha1-antitrypsin
alpha1-antitrypsin![]() antithrombin
antithrombin
Functions
The serpins, or serine proteinase inhibitors, are a superfamily of proteins that are found in a wide range of species, including plants, viruses and humans.
The family includes members as diverse as the plasma proteins alpha1-antitrypsin, alpha1-antichymotrypsin, C1q-inhibitor, antithrombin and plasminogen activator inhibitor 1, which have key regulatory functions in the inflammatory, complement, coagulation and fibrinolyttic cascades.
While most serpins control proteolytic cascades, certain serpins do not inhibit enzymes, but instead perform diverse functions such as storage (ovalbumin, in egg white), hormone carriage proteins (thyroxine-binding globulin, cortisol binding globulin) and tumor suppressor genes (maspin). The term serpin is used to describe these latter members as well, despite their noninhibitory function.
Serpins also include proteins that are involved in chromatin packing (the myeloid and erythroid nuclear termination state-specific protein or MENT), the neuron-specific protein neuroserpin and the egg-white protein ovalbumin.
Most inhibitory serpins target chymotrypsin-like serine proteases. These enzymes are defined by the presence of a nucleophilic serine residue in their catalytic site, as thrombin, trypsin and human neutrophil elastase.
Some serpins inhibit other classes of protease, as cysteine proteases, and are termed "cross class inhibitors". For example squamous cell carcinoma antigen 1 (SCCA-1) and the avian serpin myeloid and erythroid nuclear termination stage specific protein (MENT) both inhibit papain-like cysteine proteases.
The viral serpin crmA is a suppressor of the inflammatory response through inhibition of IL-1 and IL-18 processing by the cysteine protease caspase-1 (CASP1).
Extracellular roles
Approximately two thirds of human serpins perform extracellular roles. For example, extracellular serpins regulate the proteolytic cascades central to blood clotting (antithrombin), the inflammatory response (alpha1-antitrypsin, antichymotrypsin and C1q inhibitor) and tissue remodelling (PAI-1).
Non-inhibitory extracellular serpins also perform important roles. Thyroxine-binding globulin and cortisol binding globulin transport the sterol hormones thyroxine and cortisol respectively. The protease renin cleaves off a ten amino acid N-terminal peptide from angiotensinogen to produce the peptide hormone angiotensin I.
Intracellular roles
Intracellular serpins also perform roles distinct from protease inhibition. For example, maspin, a non-inhibitory serpin, is important for preventing metastasis in breast and prostate cancers. Another example is the avian nuclear cysteine protease inhibitor MENT, which acts as a chromatin remodelling molecule in avian red blood cells.
Phylogenetic studies show that most intracellular serpins belong to a single clade. Exceptions include the non-inhibitory heat shock serpin HSP47, which is a chaperone essential for proper folding of collagen and cycles between the cis-Golgi and the endoplasmic reticulum.
Cysteine proteases
Cysteine proteases differ from serine proteases in that they are defined by the presence of a nucleophilic cysteine residue, rather than a serine residue, in their catalytic site. Nonetheless, the enzymatic chemistry is similar, and serpins most likely inhibit both classes of enzyme in a similar fashion.
Structure
Members of the serpin superfamily are characterized by greater than 30% amino-acid sequence homology with alpha1-antitrypsin and have a conserved tertiary structure.
The structure of the serpins is based on three beta-sheets (A-C) and an exposed mobile reactive loop that presents a peptide sequence as a pseudosubstrate for the target proteinase.
After docking with the proteinase, the serpin undergoes a striking conformational transition that swings the proteinase from the upper to the lower pole of the serpin, in association with the insertion of an extra strand into beta-sheet A.
This transition grossly distorts the overall structure of the proteinase, thereby causing its irreversible inhibition. The serpin bound to its target enzyme is then recognized by hepatic receptors as it passes through the liver and is cleared from the circulation.
Structural biology has played a central role in the understanding of serpin function and biology. Over eighty serpin structures, in a variety of different conformations have been determined to date. Although the function of serpins varies widely, these molecules all share a common structure (or fold).
The structure of the non-inhibitory serpin ovalbumin, and the inhibitory serpin antitrypsin revealed the archetype native serpin fold.
All typically have three β-sheets (termed A, B and C) and eight or nine α-helices (hA-hI). Serpins also possess an exposed region termed the reactive centre loop (RCL) that in inhibitory molecules includes the specificity determining region and forms the initial interaction with the target protease.
In alpha1-antitrypsin, the RCL is held at the top of the molecule and is not pre-inserted into the A β-sheet. This conformation commonly exists in dynamic equilibrium with a partially inserted native conformation seen in other inhibitory serpins.
Conformational change and inhibitory mechanism
Early studies on serpins revealed that the mechanism by which these molecules inhibit target proteases appeared distinct from the lock-and-key-type mechanism utilised by small protease inhibitors such as the Kunitz-type inhibitors (eg. Basic pancreatic protease inhibitor).
Serpins form covalent complexes with target proteases. Structural studies on serpins also revealed that inhibitory members of the family undergo an unusual conformational change, termed the Stressed to Relaxed (S to R) transition.
During this structural transition the RCL inserts into β-sheet A and forms an extra (fourth) β-strand. The serpin conformational change is key to the mechanism of inhibition of target proteases.
When attacking a substrate, serine proteases catalyze peptide bond cleavage in a two-step process.
Initially, the catalytic serine performs a nucleophilic attack on the peptide bond of the substrate. This releases the new N-terminus and forms an ester-bond between the enzyme and the substrate. This covalent enzyme-substrate complex is called an acyl enzyme intermediate.
Subsequently, this ester bond is hydrolysed and the new C-terminus is released.
The RCL of a serpin acts as a substrate for its cognate protease. However, after the RCL is cleaved, but prior to hydrolysis of the acyl-enzyme intermediate, the serpin rapidly undergoes the S to R transition.
Since the RCL is still covalently attached to the protease via the ester
bond, the S to R transition causes the protease to be moved from the top to the bottom of the serpin. At the same time, the protease is distorted into a conformation where the acyl enzyme intermediate is hydrolysed extremely slowly.
The protease thus remains covalently attached to the target protease and is thereby inhibited. Further, since the serpin has to be cleaved to inhibit the target protases, inhibition consumes the serpin as well. Serpins are therefore irreversible enzyme inhibitors.
Conformational modulation of serpin activity
The conformational mobility of serpins provides a key advantage over static lock and key protease inhibitors. In particular, the function of inhibitory serpins can be readily controlled by specific cofactors.
The X-ray crystal structures of antithrombin (AT3), heparin co-factor II, MENT and murine antichymotrypsin reveal that these serpins adopt a conformation where the first two amino acids of the RCL are inserted into the top of the A β-sheet.
The partially inserted conformation is important because co-factors are able to conformationally switch partially inserted serpins into a fully expelled form. This conformational rearrangement makes the serpin a more effective inhibitor.
Certain serpins spontaneously undergo the S to R transition as part of their function, to form a conformation termed the latent state.
In latent serpins the first strand of the C-sheet has to peel off to allow full RCL insertion. Latent serpins are unable to interact with proteases and are not protease inhibitors. The transition to latency represents a control mechanism for the serpin PAI-1.
PAI-1 is released in the inhibitory conformation, however, undergoes conformational change to the latent state unless it is bound to the cofactor vitronectin. Thus PAI-1 contains an "auto-inactivation" mechanism. Similarly, antithrombin can also spontaneously convert to the latent state as part of its normal function.
Finally, the N-terminus of tengpin, a serpin from Thermoanaerobacter tengcongensis, is required to lock the molecule in the native inhibitory state. Disruption of interactions made by the N-terminal region results in spontaneous conformational change of this serpin to the latent conformation.
Serpin receptor interactions
In humans, extracellular serpin-enzyme complexes are rapidly cleared from circulation. One mechanism by which this occurs is the low density lipoprotein receptor related protein (LRP receptor), which binds to inhibitory complexes made by antithrombin, PA1-1 and neuroserpin, causing uptake and subsequent signalling events.
Thus, as a consequence of the conformational change during serpin-enzyme complex formation, serpins may act as signalling molecules that alert cells to the presence of protease activity. The fate of intracellular serpin-enzyme complexes remains to be characterised.
Conformational change and non-inhibitory function
Certain non-inhibitory serpins also use the serpin conformational change as part of their function.
For example the native (S) form of thyroxine-binding globulin has high affinity for thyroxine, whereas the cleaved (R) form has low affinity. Similarly, native (S) Cortisol Binding Globulin (CBG) has higher affinity for cortisol than its cleaved (R) counterpart.
Thus, in these serpins, RCL cleavage and the S to R transition has been commandeered to allow for ligand release, rather than protease inhibition.
Pathogenesis
The recent recognition that mutations in a serpin can also result in late-onset dementia provides insights into changes that underlie other conformational diseases, such as the amyloidoses, the prion encephalopathies and Huntington and Alzheimer diseases.
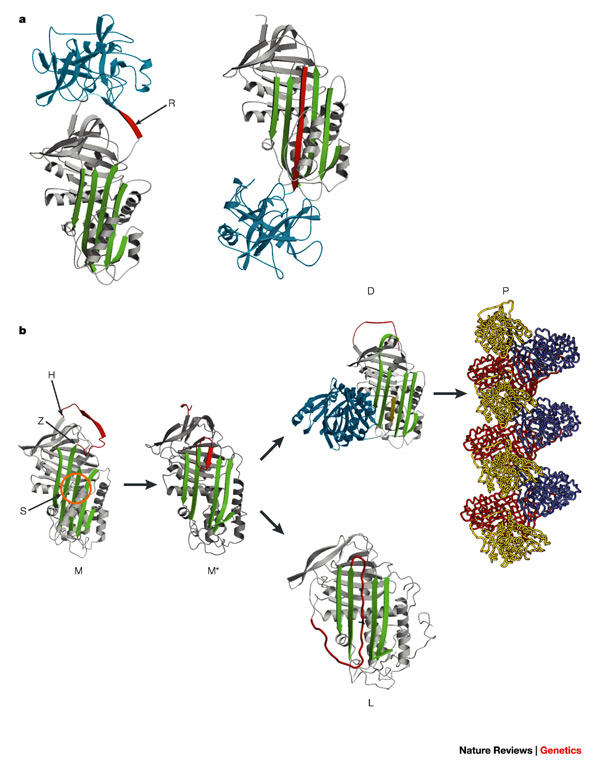
The structure of the serpins and their requirement to accept the reactive loop as an additional beta-strand is central to their role as effective antiproteinases; however, this also renders them liable to undergo conformational change that might lead to disease (the serpinopathies) (Fig. 1b).
Point mutations can destabilize beta-sheet A to allow incorporation of the reactive loop of another serpin molecule.
Sequential reactive-loop insertion results in chains of polymers that are retained in the cell of synthesis. This process is best characterized by the common condition of 1-antitrypsin deficiency in humans, which arises from the retention of this serpin in hepatocytes and is associated with liver damage.
Pathology
See: serpinopathies
The inhibitory function of the serpins involves a marked conformational transition, but this inherent molecular flexibility also renders the serpins susceptible to point mutations that result in aberrant intermolecular linkage and polymer formation.
The effects of such protein aggregation are cumulative, with a progressive loss of cellular function that results in diseases as diverse as cirrhosis and emphysema (serpinopathies).
The recent recognition that mutations in a serpin can also result in late-onset dementia provides insights into changes that underlie other conformational diseases, such as the amyloidoses, the prion encephalopathies and Huntington and Alzheimer diseases.
Members
| serpin-1 or RASP1 | serpin-2 or PI7 | serpin-B1 | serpin-B2 or PAI2 |
See also
![]() PIs
PIs![]() serpin-like proteins
serpin-like proteins![]() subtilases
subtilases![]() proprotein convertases (PCs)
proprotein convertases (PCs)![]() subtilisin-like proprotein convertases (SPCs)
subtilisin-like proprotein convertases (SPCs)
References
![]() Reichhart JM. Tip of another iceberg: Drosophila serpins. Trends Cell Biol. 2005 Dec;15(12):659-65. PMID: 16260136
Reichhart JM. Tip of another iceberg: Drosophila serpins. Trends Cell Biol. 2005 Dec;15(12):659-65. PMID: 16260136
![]() Lomas DA, Carrell RW. Serpinopathies and the conformational dementias. Nat Rev Genet. 2002 Oct;3(10):759-68. PMID: 12360234
Lomas DA, Carrell RW. Serpinopathies and the conformational dementias. Nat Rev Genet. 2002 Oct;3(10):759-68. PMID: 12360234
![]() Silverman GA, Bird PI, Carrell RW, Church FC, Coughlin PB, Gettins PG, Irving JA, Lomas DA, Luke CJ, Moyer RW, Pemberton PA, Remold-O’Donnell E, Salvesen GS, Travis J, Whisstock JC. The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. Evolution, mechanism of inhibition, novel functions, and a revised nomenclature. J Biol Chem. 2001 Sep 7;276(36):33293-6. PMID: 11435447
Silverman GA, Bird PI, Carrell RW, Church FC, Coughlin PB, Gettins PG, Irving JA, Lomas DA, Luke CJ, Moyer RW, Pemberton PA, Remold-O’Donnell E, Salvesen GS, Travis J, Whisstock JC. The serpins are an expanding superfamily of structurally similar but functionally diverse proteins. Evolution, mechanism of inhibition, novel functions, and a revised nomenclature. J Biol Chem. 2001 Sep 7;276(36):33293-6. PMID: 11435447