Notch signaling pathway
Image Gallery
[ (||image_reduire{0,60}|inserer_attribut{alt,}) ] [ (||image_reduire{0,60}|inserer_attribut{alt,}) ]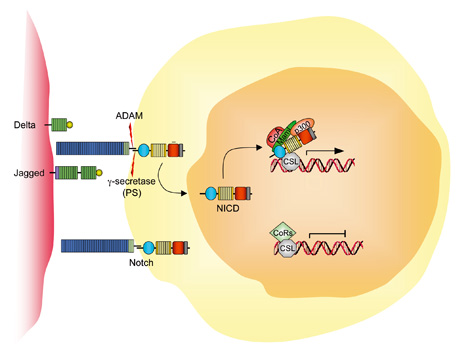{kind=link}
{kind=link}
The Notch signaling pathway is a conserved intercellular signaling mechanism that is essential for proper embryonic development in numerous metazoan organisms.
Members of the Notch gene family (NOTCHs) encode transmembrane receptors that are critical for various cell fate decisions. Multiple ligands that activate Notch and related receptors have been identified, including Serrate and Delta in Drosophila and JAG1 (MIM.601920) in vertebrates.
Four different Notch receptors (NOTCHs: NOTCH1 to NOTCH4) and five ligands (Jagged-1 (JAG1) and -2 (JAG2) and Delta-like [DLLs]: DLL1, DLL2 and DLL4) have been characterized in mammalian cells. These transmembrane receptors and ligands are expressed in different combinations in most, if not all, cell types.
The Notch pathway regulates cell fate determination of neighbouring cells through lateral inhibition, depending on their ability to express either the receptors or the ligands.
Notch family members are transmembrane receptors and developmental morphogens.
In adult tissue, activation of the NOTCH1 pathway mediates context-specific functions, such as self-renewal and T-cell differentiation. NOTCH1 signalling is initiated by the engagement of extracellular portions (ECN) of NOTCH1 with its ligands, which are members of the Jagged/Delta family.
This binding induces metalloprotease-dependent cleavage of the NOTCH1 heterodimerization domain (HD) with terminal cleavage that is dependent on gamma-secretase activity.
This process releases intracellular NOTCH1 (ICN), which translocates to the nucleus to form a multimeric transcriptional factor complex with the transcription factor CSL and co-activators of the mastermind-like (MAML) family.
These recruit additional co-activators, such as p300 and PCAF (CoA), to activate the transcription of target genes such as HES, pre-Talpha and CDKN1A. The transcriptional programmes that are initiated are thought to mediate T-cell differentiation and increase the self-renewal that is associated with NOTCH1 signalling activity.
ICN is targeted for degradation by the polyubiquitylation of sequences in its PEST domain. Mutations in the HD and PEST domain of NOTCH1 have recently been associated with T-cell acute lymphoblastic leukaemia (T-ALL).
These mutations are believed to activate NOTCH1 signalling by increasing NOTCH1 cleavage and liberation of ICN, as well as by increasing NOTCH1 protein stability by decreasing proteosomal degradation.
Pathway
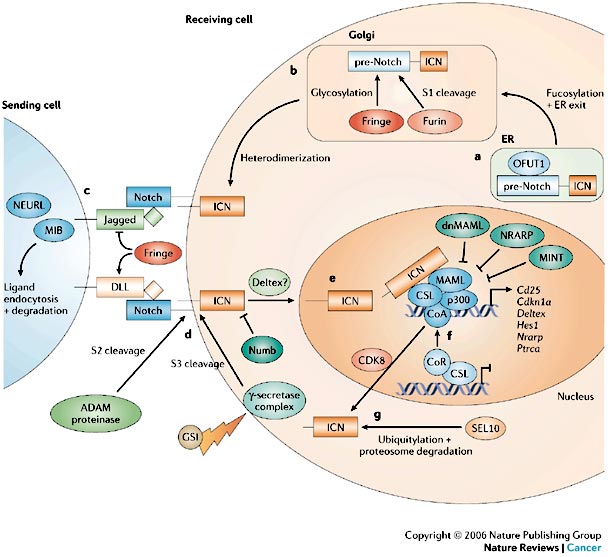
The intracellular and extracellular domains of the Notch receptor are synthesized as a single protein (pre-Notch). O-fucosyltransferase1 (OFUT1) functions as a chaperone and is required for the transport of pre-Notch from the endoplasmic reticulum (ER) to the Golgi apparatus and for fucosylation of glycosylated serine and threonine residues of the extracellular domain within the Golgi.
Glycosylation of these residues is carried out by members of the Fringe family (radical, manic and lunatic). A Furin-like convertase cleaves pre-Notch into the extracellular and intracellular domain (S1 cleavage). This results in a heterodimeric receptor with non-covalently associated domains that is transported to the plasma membrane.
The readout of receptor–ligand interaction is determined by the specific Fringe modifications that were introduced earlier in the Golgi, which affect sensitivity to the DSL (Delta (DLL)–Serrate–Lag2) ligands.
Additionally, the E3 ubiquitin ligases, Mindbomb (MIB) and Neuralized-like (NEURL) promote the turnover of the ligand and therefore contribute to productive ligand–receptor interactions.
Ligand binding initiates two successive proteolytic cleavages (S2 and S3). The first, mediated by an ADAM (a disintegrin and metalloproteinase domain) proteinase (See ADAMs), occurs in the extracellular domain.
S2 cleavage allows access of the gamma-secretase complex (GS), which is responsible for the second proteolytic cleavage (S3), which occurs within the transmembrane domain and liberates the intracellular domain (ICN).
The ICN translocates to the nucleus, where it interacts with the IPT (immunoglobulin-like fold, plexins and transcription factors)-domain-containing CSL transcription factor. Docking of the ankyrin-repeat domain of Notch to the Rel-homology region of the CSL protein generates a composite binding surface for the recruitment of Mastermind-like proteins (MAML).
These interactions convert CSL from a transcriptional repressor to an activator by displacing co-repressors (CoR) and histone deacetylases and recruiting histone acetyltransferases. MAML in turn recruits additional co-activators (CoA), such as p300.
A few transcriptional targets of Notch have been identified and include Hes1 (Hairy/enahncer of split), pre-T-cell-receptor-alpha (Ptrca), Deltex (DTX1), Nrarp (Notch-regulated ankyrin-repeat protein), Cdkn1alpha and Cd25.
Known regulators of Notch signalling include Numb, NRARP, Mint (MSX2-interacting nuclear target) and Deltex.
Numb suppresses Notch signalling, possibly by preventing nuclear localization and targeting the ICN for degradation through the E3 ligase Itch. Deltex is a positive regulator of Notch in Drosophila but can inhibit Notch signalling in mammals.
An alternative CSL-independent pathway has been described in which Notch signalling is mediated through Deltex.
The stability of nuclear ICN is regulated through its PEST (polypeptide enriched in proline, glutamate, serine and threonine) domain.
Binding of MAML to p300 and cyclin-dependent kinase 8 (CDK8) promotes hyperphosphorylation of the ICN PEST domain to facilitate ubiquitylation, probably by members of the SEL1 (also known as FBW7) family of E3 ligases and the Itch E3 ligase, which target the ICN to the proteosome.
Notch signaling
Following ligand binding, NOTCHs are activated by a series of cleavages that releases its intracellular domain (NICD).
This processing requires the activity of two proteases, namely ADAM17 (tumour necrosis factor-α converting enzyme or TACE MIM.603369) and presenilin-1 (PSEN1, MIM.104311).
Nuclear translocation of NICD results in transcriptional activation of genes of the HESs family (Hes/E(spl) family) and HEYs family (Hesr/Hey family) through interaction of NICD with RBPSUH (or CBF1 MIM.147183), Su(H), and Lag-1, which is also known as the recombination signal sequence-binding protein (RBP)-j [also called Suppressor of Hairless, Su(H)].
The specificity of each ligand for a receptor and the subsequent specificity for downstream target activation are still poorly understood. However, studies on the spatial expression of Notch components have highlighted tissue-specific functions for these proteins.
NOTCH receptors participate in a signaling pathway that regulates many aspects of morphogenesis in multicellular animals through diverse effects on differentiation, proliferation, and cell survival. Overall, when activated, Notch signalling enables neighbouring cells to acquire distinct phenotypes, through a process named lateral inhibition.
The Notch receptor is pre-cleaved in the Golgi and is targeted subsequently to the plasma membrane where it interacts with ligands located on neighbouring cells.
Receptor–ligand interaction results in a conformational change in the receptor, thus enabling additional cleavages by TACE and the γ-secretase complex.
This proteolytic activity enables the Notch intracellular domain (NICD) to translocate to the nucleus where it activates the transcription of target genes (e.g. the Hes and Hey family of transcriptional repressors).
Monoubiquitylation (Ub) of the ligand by mindbomb (MIB) induces endocytosis of the ligand and the Notch extracellular domain (NECD) into the ligand cells where additional signalling might be initiated.
Notch receptors undergo acomplex set of proteolytic processing events in response to ligand activating, which eventuallyleads to release of the intracellular domain of the receptor.Signal transduction is normally initiated by binding to transmembrane ligands of the Serrate or Delta class, which induces proteolytic release of the intracellular NOTCH domain (NICD).
Free NICD translocates to the nucleus to form a short-lived complex with a Rel-like transcription factor, CSL, and Mastermind-like co-activators that activates lineage-specific programs of gene expression.
Thus, the NOTCH pathway provides a means for local environmental cues mediated through cell:cell interactions to direct or antagonize specific developmental programs.
Notch signalling in angiogenesis
Several components of the Notch pathway are present in the vasculature, both in the endothelium (Notch1, Notch4, Jagged-1, Dll-1 and -4, Hes1, Hey1 and Hey2) and in the surrounding mural cells (which also express Notch3).
At the cellular level, Notch signalling in ECs blocks cell proliferation and favours cell survival. Inhibition of proliferation is dependent on the down-regulation of p21cip1 and the inhibition of retinoblastoma protein phosphorylation by cyclin D-cdk4.
Owing to their location, ECs are under constant aggression from inflammatory cytokines and toxins from foreign organisms. Activation of Notch signalling protects ECs from this hostile environment through activation of Bcl-2 (BCL2). Altogether, these studies suggest that Notch signalling is required for the generation of a quiescent and mature vascular phenotype.
![]() Arterial–venous differentiation
Arterial–venous differentiation
Notch signalling is also crucial for arterial–venous differentiation. This process was thought previously to be governed by environmental cues, such as blood pressure. However, recent studies demonstrate that genetic predisposition is also important.
Mutation in gridlock, the zebrafish orthologue of Hey2, results in a loss of artery-specific markers, ectopic expression of venous markers and arterial–venous shunts in mutant animals. In addition, mice with deletions in Hey1 and Hey2 have decreased arterial makers.
Finally, venous differentiation is also programmed genetically and is dependent on active repression of Notch signalling by COUP-TFII, a member of the orphan nuclear receptor superfamily.
Functions
Recent data suggest NOTCH signals control hematopoietic development at multiple decision points through effects on differentiation and self-renewal.
Components:
![]() Notch1 (NOTCH1 MIM.190198)
Notch1 (NOTCH1 MIM.190198)
![]() JAG1 (Jagged1), HES5 (MIM.607348)
JAG1 (Jagged1), HES5 (MIM.607348)
![]() MASH1 (ASCL1 MIM.100790)
MASH1 (ASCL1 MIM.100790)
![]() presenilin-2 (PSEN2 MIM.600759)
presenilin-2 (PSEN2 MIM.600759)
Pathology
![]() Vascular diseases
Vascular diseases
The role of Notch signalling in vascular development is further emphasized in two human cardiovascular diseases: the cerebral autosomal dominant arteriopathy and subcortical infarcts and leukoencephalopathy (CADASIL) and Alagille disease, which result from mutations in genes coding for Notch3 (NOTCH3) and Jagged1 (JAG1), respectively.
Additionally, over-expression of an active form of Notch4 (NOTCH4) in the vasculature results in defective embryonic angiogenesis, thus implying that crucial levels of Notch signalling are necessary for appropriate angiogenesis.
![]() In hematopoietic cells
In hematopoietic cells
Withdrawal of NOTCH signals at particular decision points leads to hypoplasia of specific lymphoid subsets, whereas enforced NOTCH signaling skews lymphoid development in the opposite direction and causes certain leukemias, including a subset of T-cell acute lymphoblastic leukemias associated with a recurrent t(7;9) translocation involving human NOTCH1.
![]() In central nervous system
In central nervous system
Notch signaling is also linked to twoforms of neurodegenerative disease. First, mutations in the Notch 3 receptor leads to the stroke anddementia syndrome CADASIL. Second, proteolytic processing of Notch is controlled by presenilins, which are frequently mutated in Alzheimer diesase.
Mouse models
In addition, mice lacking either Notch1, Jagged1 or the double knockout of Hey1 and Hey2 die during early embryonic development as a result of vascular plexus remodelling defects in the embryo and the yolk sac as well as defects in arterial venous differentiation.
Interestingly, Notch4 is expressed exclusively in the vasculature in embryonic mice but Notch4−/− mice are viable. However, Notch1 Notch4 double-knockout embryos show more pronounced vascular defects than Notch1 knockout embryos.
See also
![]() proteolyctic cleavage by notch receptors by presenilin.
proteolyctic cleavage by notch receptors by presenilin.
![]() ubiquitination and turnover of the intracellular domain of the Notch receptor
ubiquitination and turnover of the intracellular domain of the Notch receptor
References
![]() Miele L, Golde T, Osborne B. Notch signaling in cancer. Curr Mol Med. 2006 Dec;6(8):905-18. PMID: #17168741#
Miele L, Golde T, Osborne B. Notch signaling in cancer. Curr Mol Med. 2006 Dec;6(8):905-18. PMID: #17168741#
![]() High FA, Epstein JA. The multifaceted role of Notch in cardiac development and disease. Nat Rev Genet. 2008 Jan;9(1):49-61. PMID: #18071321#
High FA, Epstein JA. The multifaceted role of Notch in cardiac development and disease. Nat Rev Genet. 2008 Jan;9(1):49-61. PMID: #18071321#
![]() Rampal R, Luther KB, Haltiwanger RS. Notch signaling in normal and disease States: possible therapies related to glycosylation. Curr Mol Med. 2007 Jun;7(4):427-45. PMID: #17584081#
Rampal R, Luther KB, Haltiwanger RS. Notch signaling in normal and disease States: possible therapies related to glycosylation. Curr Mol Med. 2007 Jun;7(4):427-45. PMID: #17584081#
![]() Lasky JL, Wu H. Notch signaling, brain development, and human disease. Pediatr Res. 2005 May;57(5 Pt 2):104R-109R. PMID: #15817497#
Lasky JL, Wu H. Notch signaling, brain development, and human disease. Pediatr Res. 2005 May;57(5 Pt 2):104R-109R. PMID: #15817497#
![]() Bray SJ. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol. 2006 Sep;7(9):678-89. PMID: #16921404#
Bray SJ. Notch signalling: a simple pathway becomes complex. Nat Rev Mol Cell Biol. 2006 Sep;7(9):678-89. PMID: #16921404#
![]() Sainson RC, Harris AL. Hypoxia-regulated differentiation: let’s step it up a Notch. Trends Mol Med. 2006 Apr;12(4):141-3. PMID: #16513423#
Sainson RC, Harris AL. Hypoxia-regulated differentiation: let’s step it up a Notch. Trends Mol Med. 2006 Apr;12(4):141-3. PMID: #16513423#
![]() Kodama Y, Hijikata M, Kageyama R, Shimotohno K, Chiba T. The role of notch signaling in the development of intrahepatic bile ducts. Gastroenterology. 2004 Dec;127(6):1775-86. PMID: #15578515#
Kodama Y, Hijikata M, Kageyama R, Shimotohno K, Chiba T. The role of notch signaling in the development of intrahepatic bile ducts. Gastroenterology. 2004 Dec;127(6):1775-86. PMID: #15578515#
![]() Flynn DM, Nijjar S, Hubscher SG, de Goyet Jde V, Kelly DA, Strain AJ, Crosby HA. The role of Notch receptor expression in bile duct development and disease. J Pathol. 2004 Sep;204(1):55-64. PMID: #15307138#
Flynn DM, Nijjar S, Hubscher SG, de Goyet Jde V, Kelly DA, Strain AJ, Crosby HA. The role of Notch receptor expression in bile duct development and disease. J Pathol. 2004 Sep;204(1):55-64. PMID: #15307138#
![]() Kadesch T. Notch signaling: the demise of elegant simplicity. Curr Opin Genet Dev. 2004 Oct;14(5):506-12. PMID: #15380241#
Kadesch T. Notch signaling: the demise of elegant simplicity. Curr Opin Genet Dev. 2004 Oct;14(5):506-12. PMID: #15380241#
![]() Weng AP, Aster JC. Multiple niches for Notch in cancer: context is everything. Curr Opin Genet Dev. 2004 Feb;14(1):48-54. PMID: #15108805#
Weng AP, Aster JC. Multiple niches for Notch in cancer: context is everything. Curr Opin Genet Dev. 2004 Feb;14(1):48-54. PMID: #15108805#
![]() Radtke F, Raj K. The role of Notch in tumorigenesis: oncogene or tumour suppressor? Nat Rev Cancer. 2003 Oct;3(10):756-67. PMID: #14570040#
Radtke F, Raj K. The role of Notch in tumorigenesis: oncogene or tumour suppressor? Nat Rev Cancer. 2003 Oct;3(10):756-67. PMID: #14570040#
![]() Miyamoto Y, Maitra A, Ghosh B, Zechner U, Argani P, Iacobuzio-Donahue CA, Sriuranpong V, Iso T, Meszoely IM, Wolfe MS, Hruban RH, Ball DW, Schmid RM, Leach SD. Notch mediates TGF alpha-induced changes in epithelial differentiation during pancreatic tumorigenesis. Cancer Cell. 2003 Jun;3(6):565-76. PMID: #12842085#
Miyamoto Y, Maitra A, Ghosh B, Zechner U, Argani P, Iacobuzio-Donahue CA, Sriuranpong V, Iso T, Meszoely IM, Wolfe MS, Hruban RH, Ball DW, Schmid RM, Leach SD. Notch mediates TGF alpha-induced changes in epithelial differentiation during pancreatic tumorigenesis. Cancer Cell. 2003 Jun;3(6):565-76. PMID: #12842085#
![]() Gridley T. Notch signaling and inherited disease syndromes.
Hum Mol Genet. 2003 Apr 1;12 Spec No 1:R9-13. PMID: #12668592#
Gridley T. Notch signaling and inherited disease syndromes.
Hum Mol Genet. 2003 Apr 1;12 Spec No 1:R9-13. PMID: #12668592#
![]() Screpanti I, Bellavia D, Campese AF, Frati L, Gulino A. Notch, a unifying target in T-cell acute lymphoblastic leukemia? Trends Mol Med. 2003 Jan;9(1):30-5. PMID: #12524208#
Screpanti I, Bellavia D, Campese AF, Frati L, Gulino A. Notch, a unifying target in T-cell acute lymphoblastic leukemia? Trends Mol Med. 2003 Jan;9(1):30-5. PMID: #12524208#
![]() Joutel A, Tournier-Lasserve E. Notch signalling pathway and human diseases. Semin Cell Dev Biol. 1998 Dec;9(6):619-25. PMID: #10075489#
Joutel A, Tournier-Lasserve E. Notch signalling pathway and human diseases. Semin Cell Dev Biol. 1998 Dec;9(6):619-25. PMID: #10075489#