Home > D. General pathology > Blood and immunity > Dysimmune diseases > allograft rejection
allograft rejection
Monday 15 March 2004
Digital case
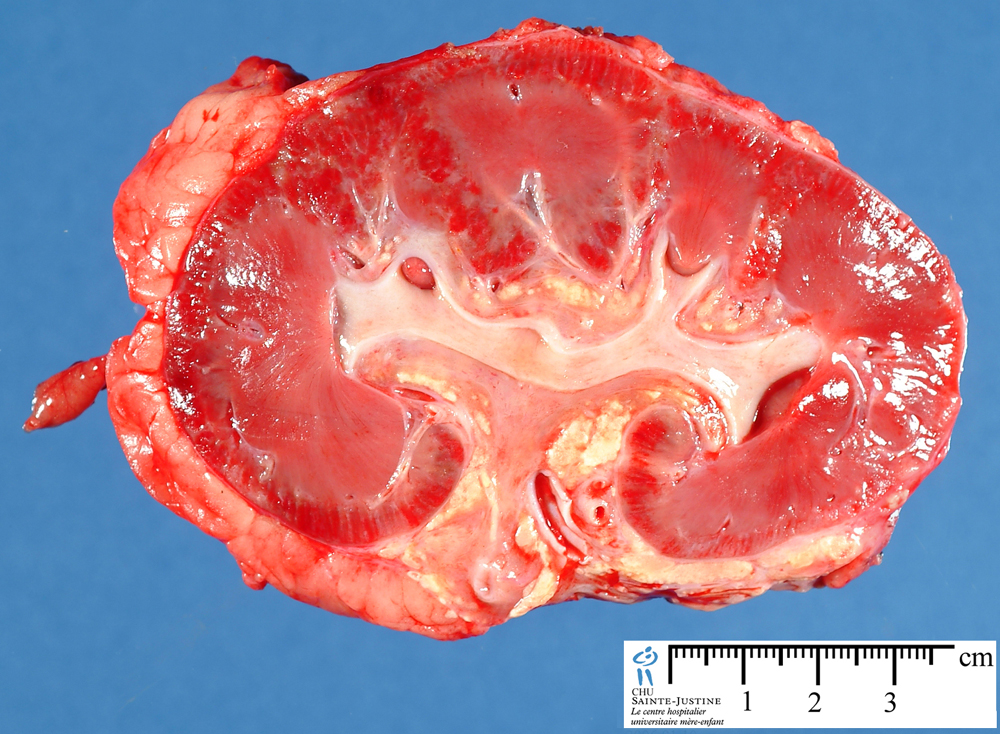
UI:774 : acute renal allograft rejection
Transplant rejection involves several of the immunologic reactions. A major barrier to transplantation is the process of rejection, in which the recipient’s immune system recognizes the graft as being foreign and attacks it. One of the important goals of present-day immunologic research is successful transplantation of tissues in humans without rejection.
Although the surgical expertise for the transplantation of skin, kidneys, heart, lungs, liver, spleen, bone marrow, and endocrine organs is now well in hand, it outpaces thus far the ability to confer on the recipient permanent acceptance of foreign grafts.
Mechanisms Involved in Rejection of Kidney Grafts
As stated above, graft rejection depends on recognition by the host of the grafted tissue as foreign. The antigens responsible for such rejection in humans are those of the HLA system. Because HLA genes are highly polymorphic, any two individuals (other than identical twins) will express some HLA proteins that are different.
Thus, every individual will recognize some HLA molecules in another individual as foreign (allogeneic) and will react against these. This reaction is the basis of rejection of grafts from one individual to another.
Rejection is a complex process in which both cell-mediated immunity and circulating antibodies play a role; moreover, the relative contributions of these two mechanisms to rejection vary among grafts and are often reflected in the histologic features of the rejected organs.
T Cell-mediated rejection
The critical role of T cells in transplant rejection has been documented both in humans and in experimental animals. T cell-mediated graft rejection is called cellular rejection, and it is induced by two mechanisms: destruction of graft cells by CD8+ CTLs and delayed hypersensitivity reactions triggered by activated CD4+ helper cells.
The recipient’s T cells recognize antigens in the graft (the allogeneic antigens, or alloantigens) by two pathways, called direct and indirect.
In the direct pathway, T cells of the transplant recipient recognize allogeneic (donor) MHC molecules on the surface of an antigen-presenting cell in the graft.
It is believed that dendritic cells carried in the donor organs are the most important immunogens because they not only richly express class I and II HLA molecules but also are endowed with costimulatory molecules (e.g., B7-1 and B7-2).
The T cells of the host encounter the dendritic cells either within the grafted organ or after the dendritic cells migrate to the draining lymph nodes.
Both the CD4+ and the CD8+ T cells of the transplant recipient are involved in this reaction. CD8+ T cells recognize class I HLA antigens and differentiate into mature CTLs.
This process of CTL differentiation is complex and incompletely understood. It appears to be dependent on the release of cytokines, such as IL-2, from CD4+ helper cells and CD40 ligand on the helper cells activating antigen-presenting cells to promote the differentiation of CTLs. Once mature CTLs are generated, they kill the grafted tissue by mechanisms already discussed.
The CD4+ helper T-cell subset is triggered into proliferation and differentiation into TH1 effector cells by recognition of allogeneic class II molecules. As in delayed hypersensitivity reactions, cytokines secreted by the activated CD4+ T cells cause increased vascular permeability and local accumulation of mononuclear cells (lymphocytes and macrophages), and activate the macrophages, resulting in graft injury.
The direct recognition of allogeneic MHC molecules seems paradoxical to the rules of self-MHC restriction: If T cells are normally restricted to recognizing foreign peptides displayed by self-MHC molecules, why should these T cells recognize foreign MHC?
Such recognition has been explained by assuming that allogeneic MHC molecules, with their bound peptides, resemble, or mimic, the self-MHC-foreign peptide complexes that are recognized by self-MHC-restricted T cells. The structural basis of such mimicry is not entirely clear.
In the so-called indirect pathway of allorecognition, recipient T lymphocytes recognize antigens of the graft donor after they are presented by the recipient’s own antigen-presenting cells.
This process involves the uptake and processing of MHC molecules from the grafted organ by host antigen-presenting cells. The peptides derived from the donor tissue are presented by the host’s own MHC molecules, like any other foreign peptides.
Thus, the indirect pathway is similar to the physiologic processing and presentation of other foreign (e.g., microbial) antigens. The indirect pathway generates CD4+ T cells that enter the graft and recognize graft antigens being displayed by host antigen-presenting cells that have also entered the graft, and the result is a delayed hypersensitivity type of reaction.
However, CD8+ CTLs that may be generated by the indirect pathway cannot directly recognize or kill graft cells, because these CTLs recognize graft antigens presented by the host’s antigen-presenting cells. Therefore, when T cells react to a graft by the indirect pathway, the principal mechanism of cellular rejection may be T-cell cytokine production and delayed hypersensitivity.
It is postulated that the direct pathway is the major pathway in acute cellular rejection, whereas the indirect pathway is more important in chronic rejection. However, this separation is by no means absolute.
Antibody-mediated rejection
Although there is little doubt that T cells are pivotal in the rejection of organ transplants, antibodies evoked against alloantigens in the graft can also mediate rejection. This process is called humoral rejection (antibody-mediated rejection), and it can take two forms. Hyperacute rejection occurs when preformed antidonor antibodies are present in the circulation of the recipient.
Such antibodies may be present in a recipient who has already rejected a kidney transplant. Multiparous women who develop anti-HLA antibodies against paternal antigens shed from the fetus may also have preformed antibodies to grafts taken from their husbands or children, or even from unrelated individuals who share HLA alleles with the husbands.
Prior blood transfusions can also lead to presensitization because platelets and white blood cells are rich in HLA antigens and donors and recipients are usually not HLA-identical. When preformed antidonor antibodies are present, rejection occurs immediately after transplantation because the circulating antibodies react with and deposit rapidly on the vascular endothelium of the grafted organ.
Complement fixation occurs, resulting in thrombosis of vessels in the graft, and ischemic death of the graft. With the current practice of cross-matching, that is, testing recipient’s serum for antibodies against donor’s cells, hyperacute rejection is no longer a significant clinical problem.
In recipients not previously sensitized to transplantation antigens, exposure to the class I and class II HLA antigens of the donor may evoke antibodies. The antibodies formed by the recipient may cause injury by several mechanisms, including complement-dependent cytotoxicity, inflammation, and antibody-dependent cell-mediated cytotoxicity.
The initial target of these antibodies in rejection appears to be the graft vasculature. Thus, antibody-dependent, or acute humoral rejection, is usually manifested by a vasculitis, sometimes referred to as rejection vasculitis.
Morphology of Rejection Reactions. On the basis of the morphology and the underlying mechanism, rejection reactions are classified as hyperacute rejection, acute rejection, and chronic rejection.
Types
hyperacute rejection
acute rejection
- humoral acute rejection
- cellular acute rejection
chronic rejection
- chronic humoral rejection (antibody-mediated rejection)
According to the localization
cardiac allograft rejection
hepatic allograft rejection
renal allograft rejection
cutaneous allograft rejection (skin allograft rejection)